Tilbake til artikkelserien om metabolismen
Fettvevet er kroppens primære energilager, og sammen med glukose er fettsyrer en viktig energikilde. For å frigi energi må fettsyrene brytes ned, og i denne artikkelen ser vi nærmere på hvordan og hvor dette foregår.
Denne artikkelen tilhører artikkelserien om metabolismen
Relevante artikler
Fettsyrenedbrytning
Fettsyrer består av lange kjeder karbonatomer med en karboksylsyregruppe i den ene enden, og disse kommer i ulike varianter. Vi har mettede fettsyrer, der alle karbonatomene er bundet sammen med enkeltbindinger, og vi har umettede fettsyrer som inneholder en eller flere dobbeltbindinger i karbonkjeden. Fettsyrene kommer også i ulike lengder. Lagrings- og transportformen for fettsyrer er primært som triglyserider, som består av tre fettsyrer bundet til et glyserolmolekyl. Triglyseridene kan ikke krysse cellemembraner, så fettsyrene må spaltes av fra glyserolmolekylet ved hjelp av en lipase. Vi har ulike lipaser som gjennomfører dette, eksempelvis lipoprotein lipase (LPL) som sørger for opptak av fettsyrer til cellene fra lipoproteiner, og hormonsensitiv lipase (HSL) som frigjør fettsyrer fra triglyseridene i fettvevet for eksport til blodet. Glyserol kan etter spaltingen metaboliseres videre f.eks. til i glukoneogenese/glykolyse eller brukes for å danne nye triglyserider.
Fettsyrer kan brukes som energi i nesten alle cellene i kroppen, og forbrenningen av disse skjer i mitokondriene. De cellene som ikke har mitokondrier kan dermed ikke bruke fettsyrer direkte som energikilde. Mesteparten av kroppens fettsyreoksidasjon foregår i leveren og frigjør energi.
Før en fettsyre kan oksideres må den aktiveres ved å kobles til koenzym-A (CoA). Dette skjer ved hjelp av enzymet acyl-CoA syntetase som befinner seg i mitokondrienes ytre membran, og er en energikrevende prosess. For å fraktes inn i selve mitokondriet, der β-oksidasjonen foregår, kobles fettsyren til et karnitinmolekyl ved hjelp av enzymet karnitin palmitoyl transferase 1 (CPT1), som også befinner seg i den ytre mitokondrielle membranen. Et spesialisert transportsystem frakter dermed acylkarnitin inn i mitokondriet, og på innsiden byttes karnitinmolekylet igjen med CoA ved hjelp av karnitin palitoyl acyltransferase II (CPT2) slik at vi nå har en fettsyre-CoA inne i mitokondriet klar for å gjennomgå β-oksidasjon. Mellomkjedede fettsyrer trenger ikke å kobles til karnitin for å fraktes inn i mitokondriet.
β-oksidasjonen
Selve nedbrytningen av fettsyrer kalles beta(β)-oksidasjon, et navn som kommer av at reaksjonen skjer på det andre karbonatomet (betakarbonet) i fettsyrekjeden, ikke inkludert karbonatomet i karboksylsyregruppen. Resultatet av en runde med β-oksidasjon er at det spaltes av en tokarbonenhet (acetyl-CoA) fra fettsyren. I tillegg dannes det et hver av de elektronbærende molekylene FADH2 (riboflavin. Produksjonen av ATP, som er kroppens energimolekyl, foregår hovedsakelig i disse prosessene. Bildet under viser en skjematisk oversikt over β-oksidasjon.
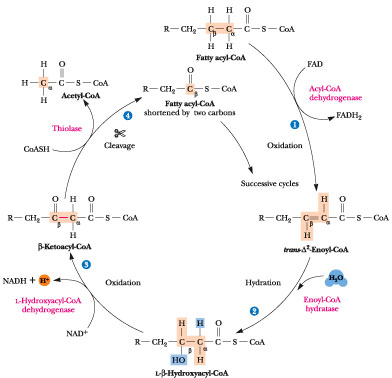
β-oksidasjon av mettede fettsyrer foregår i fire enzymatiske steg:
Det første steget er en oksideringsreaksjon der fettsyre-CoA ved hjelp av en acyl-CoA dehydrogenase blir omdannet til enoyl-CoA, som inneholder en dobbeltbinding mellom karbon nr 2 og 3. I dette steget får vi dannet det elektronbærende molekylet FADH2.
I det neste steget kobles det et vannmolekyl til denne dobbeltbindingen, og vi får dannet et hydroksyacyl-CoA. Denne prosessen katalyseres av enoyl CoA hydratase. som du ser på figuren så forsvinner dobbeltbindingen som ble dannet i det første trinnet.
Den tredje reaksjonen er en ny oksidasjonsreaksjon, hvor hydroksyacyl-CoA blir omdannet til et ketoacyl-CoA. Reaksjonen er katalysert av L-3-hydroksyacyl CoA dehydrogenase. I denne prosessen får vi dannet det elektronbærende molekylet NADH.
Det siste trinnet i β-oksidasjonen er en kløyvingsreaksjon der vi får spaltet av acetyl-CoA, som kan metaboliseres videre i sitronsyresyklusen. Vi står da igjen med en fettsyre-CoA som er to karboner kortere, som kan gå inn i en ny runde med β-oksidasjon.
Umettede fettsyrer må mettes før de kan oksideres på denne måten, og dette skjer ved hjelp av to ekstra enzymatiske steg. Grunnen til at dette er nødvendig er at dobbeltbindingen hindrer acyl-CoA dehydrogenase som katalyserer det første steget i β-oksidasjonen. Vi har derfor to enzymer, enoyl-CoA isomerase og 2,4-dienoyl-CoA reduktase, som bearbeider denne dobbeltbindingen og gjør at vi sitter igjen med et substrat for β-oksidasjon.
Fettsyrer med oddetall karbonatomer kan ikke oksideres fullstendig ved β-oksidasjon. Den siste runden vil produsere en fettsyre-CoA med tre karbonatomer, propionyl-CoA, og denne må metaboliseres på en annen måte. Propionyl-CoA karboksyleres først ved hjelp av enzymet propionyl-CoA karboksylase som bruker biotin som kofaktor. Succinyl-CoA er et intermediat i sitronsyresyklusen og går inn i denne.
Peroksisomal β-oksidasjon
Peroksisomene er noen små organeller inni cellene som inneholder en rekke enzymer som blant annet er involvert i nebrytningen av fettsyrer. Selv om mesteparten av fettsyrenedbrytningen skjer i mitokondriene, må lange fettsyrer må først forkortes i peroksisomene for å være et godt substrat for mitokondriell β-oksidasjon. Endeproduktet for peroksisomal β-oksidasjon er oktanoyl-CoA, som har 8 karbonatomer i fettsyrekjeden, og fettsyrer som er kortere enn dette kan bare forbrennes i mitokondriene.
β-oksidasjon i peroksisomene skiller seg fra oksidasjonen i mitokondriene ved den første oksideringsreaksjonen. I denne reaksjonen fikk vi i mitokondriene produsert det elektronbærende molekylet FADH2, men i peroksisomene får vi i stedet dannet hydrogenperoksid, som er reaktiv oksygenforbindelse. Dette skyldes at peroksisomene ikke er direkte koblet til elektrontransportkjeden, og elektronene som frigjøres blir derfor omdannet til hydrogenperoksid i stedet. Peroksisomene har imidlertid også rikelig av et enzym som heter katalase, som degraderer hydrogenperoksid slik at det dannes vann. De resterende trinnene av den peroksisomale β-oksidasjonen er identisk til det som foregår i mitokondriene.
Noen fettsyrer er forgrenede, og disse kan ikke brytes ned ved β-oksidasjon. Disse fraktes derfor inn i peroksisomene hvor vi finner en α-hydroksylase. Denne kan bryte ned denne forgreningen i en prosess som kalles α-oksidasjon. Etter at forgreningen er brutt ned kan resten av fettsyren oksideres på vanlig måte.
Artikkelen er sist oppdatert august 2013