Tilbake til artikkelserien om menneskets fysiologi
Som oftest snakker vi om hjertet i entall, til tross for det er to separate pumper som utgjør organet. Den høyre pumpen, eller halvdelen, pumper blod gjennom lungene, og den venstre halvdelen pumper blodet gjennom de perifere vevene, eller resten av kroppen. Både høyre og venstre pumpe består av to deler, atriumet og ventrikkelen. Atriumet i hver halvdel pumper blod til ventrikkelen, mens ventrikkelen i tur tar seg av den mest krevende delen av jobben som er å pumpe blodet videre til lungene (pulmonær sirkulasjon) eller resten av kroppen (perifér sirkulasjon). Dette skjer ved at hjertemuskelen kontraherer ved hjelp av aksjonpotensial som eksiterer myocyttene (muskelcellene) og skaper kontraksjon. Hjertets ledningssystem vil jeg komme tilbake til i detalj i en av de neste artiklene, selv om noe av det vil diskuteres også i denne artikkelen ettersom hjertets aksjonspotensial vil bli beskrevet her.
Hjertemuskelens anatomi
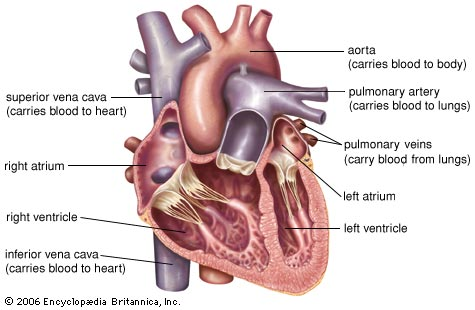
Hjertemuskelen har flere likhetstrekk med skjelettmuskulatur og inneholder mye kontraktilt muskelvev som aktin og myosin. Til tross for likhetene er hjertemuskulaturen, som vi skal se under, også nokså ulik vanlig muskulatur. Viktige strukturelle elementer som de ulike hjertekamrene (atriene og ventriklene) samt hjerteklaffene (trikuspidal-, mitral-, aorta- og pulmonalklaffen) vil bli diskutert senere i artikkelen, men under følger likevel en kort oversikt over hjerteposen og hjerteveggens anatomi.
Perikardet
Hjertet er plassert i et væskefylt hulrom som kalles det perikardiale hulrommet. Veggene i dette hulrommet utgjøres av en spesiell membran og kalles perikardet, eller hjerteposen. Membranen i hjerteposen produserer en serøs væske som smører hjertet og forhindrer friksjon mellom hjertet og andre organer. I tillegg holder hjerteposen hjertet på plass samtidig som det skaper et hulrom som tillater hjertet å ekspandere når det fylles med blod. Hjerteposen består av to lag, hvorav et visceralt lag som dekker utsiden av hjertet, og et parietalt lag som dekker utsiden av det perikardiale hulrommet.
Hjerteveggen
Hjerteveggen består av 3 lag; epikardiet, myokardiet og endokardiet. Epikardiet er det ytterste laget av hjerteveggen, og er egentlig bare et annet ord for det viscerale laget til hjerteposen, og har dermed de samme funksjonene, som smøring og produksjon av serøs væske. Mykoardiet er det midterste og tykkeste laget av hjerteveggen og består av selve hjertemuskelen som er ansvarlig for å pumpe blodet rundt i kroppen vår. Innerst i hjerteveggen finner vi endokardiet, som er et enkelt endotellag som dekker innsiden av hjertet, og forhindrer blodet i å klistre seg til innsiden av hjertet.
Tykkelsen på hjerteveggen varierer mellom de ulike delene av hjertet. I atriene, som det står mer om under, vil hjerteveggen være langt tynnere, ettersom de ikke skal pumpe blod lenger enn ned i ventriklene. Ventriklene har derimot en mye tykkere hjertevegg, ettersom det trengs langt mer kraft for å pumpe blod til lungene og resten av kroppen. Venstre ventrikkelhalvdel er større enn høyre, da denne halvdelen er ansvarlig for å pumpe blod til hele kroppen i motsetning til høyre som pumper til lungene.
Hjertemuskelens Fysiologi
Hjertet er satt sammen av tre hovedmuskelfibre; 1)atriale, 2) ventrikulære og 3) spesialiserte eksitatoriske og “strømledende” muskelfibre. De atriale og ventrikulære muskelfibrene kontraherer (trekker seg sammen) slik som vanlig skjelettmuskulatur, med det untaket at kontraksjonen varer mye lenger. I motsetning vil de eksitatoriske og strømledende muskelfibrene utøve en mye svakere kontraksjon ettersom de inneholder få kontraktile muskelfibre. I stedet vil hovedfunksjonen deres være å lede aksjonspotensialer gjennom hjertet, som skaper det karakteristiske, rytmiske hjerteslaget.
Spesifikk anatomi
Hjertemuskelfibre utformes av flere individuelle men sammenkoblede celler som ligger paralellt med hverandre. Cellene sammenkobles ved at cellemembranene til hver individuelle celle fusjonerer med hverandre, og dette tillater i tur rask utvekskling av ioner som kan karakteriseres som en form for kommunikasjon mellom celler man ikke finner i andre vev. Når cellene ligger sammenkoblet på denne måten kalles de for et syncytium (se bilde under). På den måten vil aksjonspotensialene flyte med letthet fra den ene hjertemuskelcellen til den andre. Det skal ikke mer enn èn eksitasjon til før aksjonspotensialet spres til alle cellene.
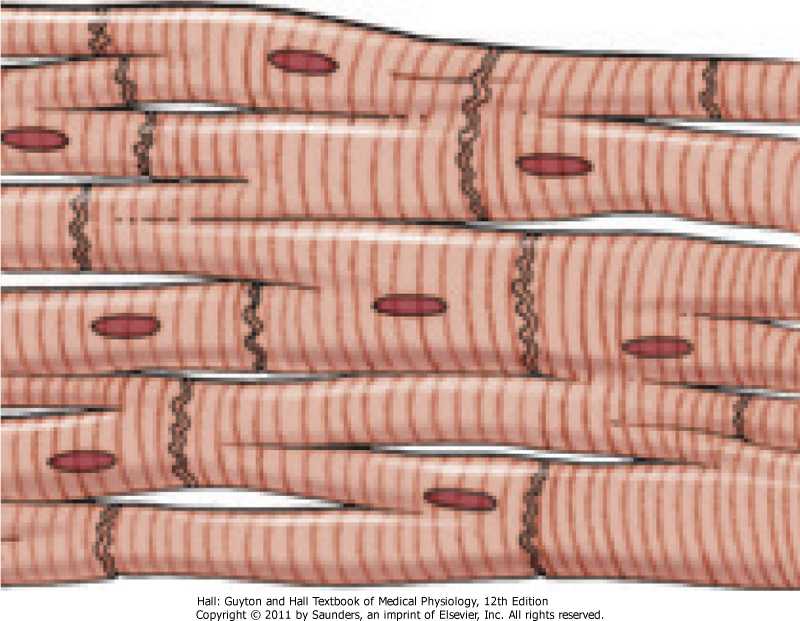
Hjertet er sammensatt av to slike syncytier, det atriale og det ventrikulære som henholdsvis utgjør veggen på de to atriene og ventriklene. Atriene er separert fra de to ventriklene av et fibrøst lag som dekker den atrioventrikulære åpningen. På grunn av det fibrøse laget som dekker atriventrikulære åpningen, ledes ikke aksjonspotensialene direkte fra det atrielle syncytiet til det ventrikulære syncytiet. I stedet vil denne ledningen gå via den atrioventrikulære knuten, også kalt AV-knuten, som er et knutepunkt bestående av flerfoldige strømledende fibre. Hjertets ledningssystem vil bli diskutert i større detalj i senere.
Oppdelingen av hjertet i to separate, funksjonelle syncytier gjør at atriet kontraherer litt tidligere enn ventrikkelen. Dette er svært viktig for effektiv pumping av blod rundt i kroppen.
Hjertets aksjonspotensial
Aksjonspotensialet til hjertemuskelfibrene som er vist på figuren under, har en styrke på gjennomnsittlig 105 millivolt, som betyr at det intracellulære potensialet stiger fra en negativ verdi på ca -85 millivolt til ca +20 milivolt ved hvert hjerteslag. Etter den første “toppen” forblir membranen depolarisert i ca 0.2 sekunder, og når dermed et såkalt platå som følges av en brå repolarisering. Dette platået i aksjonspotensialet gjør at hjertemuskelen kontraherer over mye lenger tid (opp til 15 ganger lenger) enn et vanlig skjelettmuskelfiber.
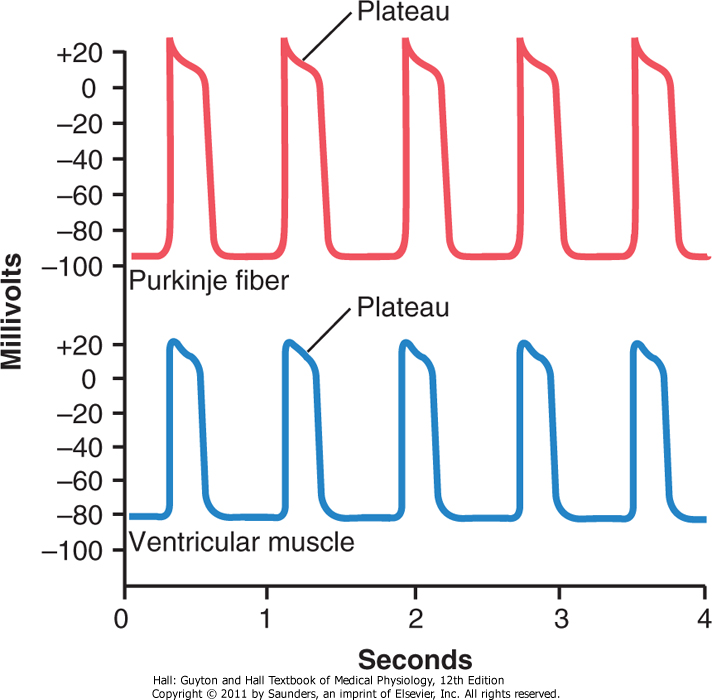
Årsaken til forskjellen på lengden av aksjonspotensialet samt platået som oppstår skyldes to ting. 1) Aksjonspotensialet i muskelceller initieres ved at såkalte “raske” natriumkanaler åpnes og lukkes i løpet av noen tusendeler av et sekund, slik at både depolarisering og repolarisering skjer ekstremt hurtig. I hjertemuskulatur åpnes to typer ionekanaler deriblant de samme, raske natriumkanalene man finner i skjelettmuskulatur, men også “trege” kalsiumkanaler. Disse kalsiumkanalene åpnes saktere, men forblir åpne mye lenger enn natriumkanalene, som fører til lenger depolariseringsperiode og i tur platået som oppstår. Kalsiumet som strømmer inn i hjertemuskelcellene fører deretter til at muskelfibrene trekker seg sammen, mens i skjelettmuskelcellene kommer kalsiumet fra sarkoplasmatisk retikulum ettersom muskelceller ikke har disse kalsiumkanalene. 2) Umiddelbart etter starten av aksjonpotensialet reduseres permeabiliteten for kalium-ioner i hjertemuskelcellene, slik at disse ikke lekker ut like hurtig som i skjelettmuskulatur og repolarisererer membranen. Etter 0.2-0.3 sekunder når de trege kalsiumkanalene lukkes og innstrømningen av både kalsium og natrium stoppes, økes også kaliumpermeabiliteten kraftig og kalium vil strømme ut av cellen og repolariserer membranen, og aksjonspotensialet stopper opp.
Refraktærperioden
Med refraktærperioden menes det tidsrommet hvor det ikke kan gå aksjonpotensial i hjertet. Dette skyldes at det allerede går et aksjonspotensial, og hjertecellene kan ikke re-eksiteres når dette skjer. Dette kalles den absolutte refraktærperioden. En relativt refraktærperiode følger umiddelbart etter repolariseringen av cellene. Da er det mulig at et nytt aksjonspotensial kan oppstå, men avhenger av et kraftigere stimuli enn normalt. Refraktærperioden til atriene er langt kortere enn i ventriklene.
Eksitasjons-kontraksjonkobling
Eksitasjons-kontraksjonskobling beskriver hvordan aksjonspotensialet får muskelfibrene til å kontrahere. Når et aksjonpotensial går, spres dette inn i dypet av hjertemuskelen ved hjelp av tverrgående tubuli (T-rør) som går tvers igjennom hjertemuskelcellene og dermed kommuniserer med både ekstracellulærvæsken, og innsiden av cellen. Disse T-rørene interagerer også med langsgående sarkoplasmatiske tubuli som dermed fører til frigjøring av kalsium fra sarkoplasmatisk retikulum inni hjertemuskelcellen, som i tur utløser den kjemiske reaksjonen som oppstår når musklene trekker seg sammen.
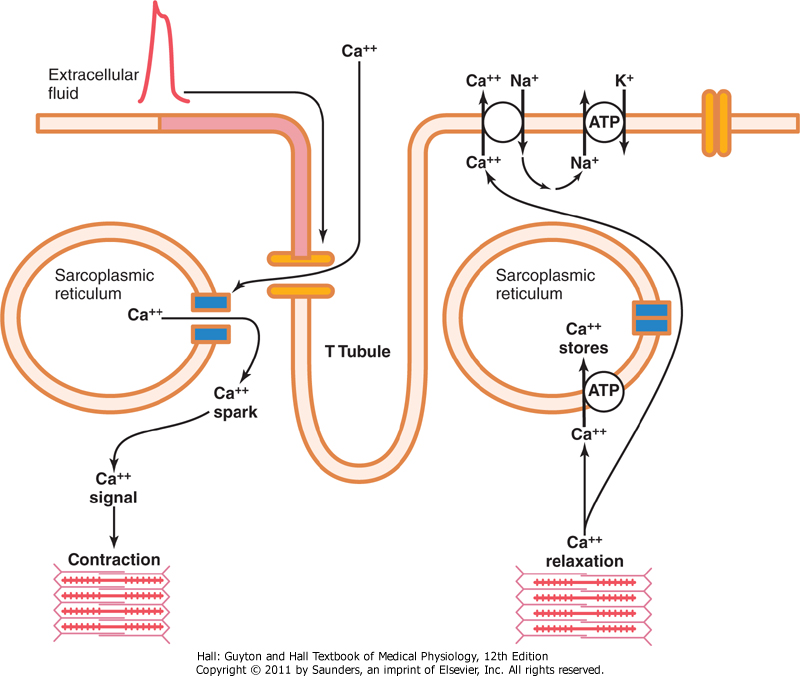
Så langt er det små forskjeller på ekstitasjons-kontraksjonkobling mellom hjertemuskelceller og skjelettmuskelceller da det samme skjer her. Men det er èn vesentlig forskjell, og det er at T-rørene i hjertet også er et lager for kalsiumioner. Slik at når aksjonspotensialet ledes langs T-rørene frisgjøres det kalsium som diffunderer inn i cellen, binder seg til reseptorer i det sarkoplasmatiske retikulum som øker kalsiumfrigjøring i hjertemuskelcellen ytterligere. Dette fører så til at flere kalsiumkanaler åpnes i T-rørene, og enda mer kalsium strømmer inn i cellen og skaper kontraksjon som i en vanlig skjelettmuskelcelle. Uten denne ekstra kalsium-frigjøringen fra T-rørene ville kontraksjonen været vesentlig svakere, og hjertet ville ikke klart å pumpe med samme kraft. Dette skyldes at hjertemuskelcellenes sarkoplasmatiske retikulum ikke er like velutviklet som i skjellettmuskelceller, og lagrer ikke nok kalsium til å stimulere en kraftig kontraksjon alene. I motsetning er hjertets T-rør 5 ganger større en T-rør i skjelettmuskelcellene, og kan i tillegg lagre kalsium, noe T-rør i skjelettmuskelceller som nevnt ikke gjør.
I avslutningsfasen av hjertets aksjonspotensial stopper innstrømningen av kalsium samtidig som kalsiumet som allerede befinner seg i sarkoplasma pumpes effektivt tilbake i sarkoplasmatisk retikulum såvel som til T-rørene. Dette skjer ved hjelp av en kalsium-ATPase pumpe. Kalsiumet kan i tillegg pumpes ut ved hjelp av en natrium-kalsium kanal, hvor intracellulært kalsium “byttes” mot ekstracellulært natrium. Dette natriumet pumpes så også via natrium-kalium pumpen. Dette resulterer i at kontraksjonen stopper opp før et nytt aksjonspotensial blir fyrt av.
Hjertesyklusen
Hjertesyklusen betegner alle hendelsene som finner sted i hjertet fra begynnelsen av et hjerteslag. Hver syklus begynner med en spontan eksitasjon av sinusknuten (mer om denne i neste del) som vi finner i høyre atrie, nær åpningen av øvre vena cava, og aksjonspotensialet ledes raskt herfra gjennom begge atrier, gjennom den atrioventrikulære knuten (AV-knuten) og ned i ventrikkelen. Grunnet ledningssystemets spesielle oppbygning tar det mer enn 0.1 sekund før aksjonspotensialet har nådd ventriklene. Som nevnt tidligere gjør dette at atriene kontraherer før ventriklene, noe som tillater pumping av blod fra atriene til ventriklene før ventriklene kontraherer langt kraftigere for å frakte blodet til lungene og rundt ellers i kroppen.
Diastole og systole
Hjertesyklusen består av en hvileperiode, diastole, hvor hjertet fylles med blod, og en arbeidsperiode, systole, hvor hjertet pumper ut blod.
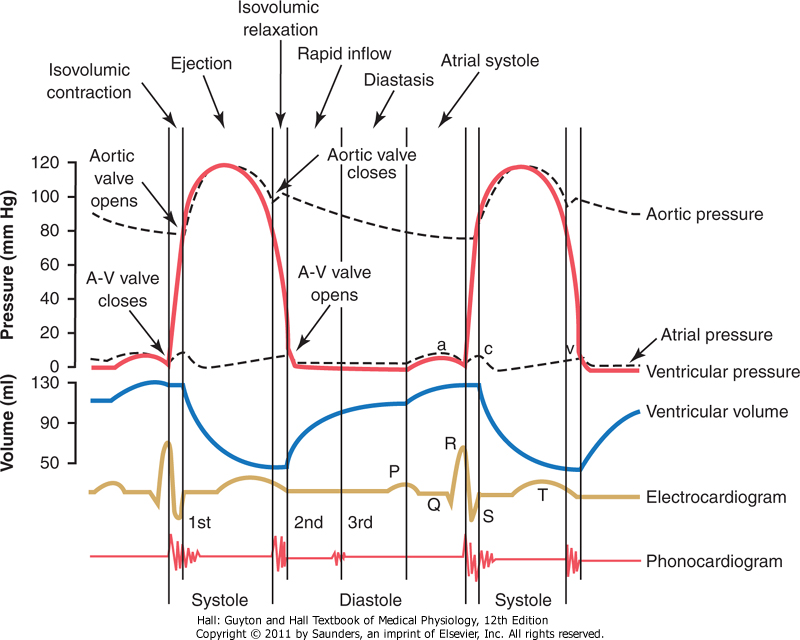
Figuren over viser de ulike hendelsene som finner sted i hjertesyklusen for venstre halvdel. De tre øverste kurvene viser henholdsvis trykk-forandringene i aorta, venstre ventrikkel og venstre atrium. Den fjerde kurven viser forandringer i blodvolumet i venstre ventrikkel, den femte et elektrokardiogram (EKG), og den sjette et fonokardiogram som beskriver lyden fra hjerteklaffene.
Atriene
Blodet tømmes normalt sett i atriene fra de store venene. 80% av blodet strømmer direkte ned i ventriklene før atriene kontraherer, før kontraksjonen sørger for at de resterende 20% blir pumpet ned i ventrikkelen. På den måten fungerer atriene som grunnpumper, som øker ventriklenes effektivitet med så mye som 20%. På den andre siden vil hjertet helt fint kunne fungere uten atriefunksjon, ettersom det er laget for å pumpe 3-400 ganger mer blod enn det som trengs i hvile. Mangelen på atriefunksjon merkes derfor gjerne ikke før tung fysisk aktivitet, hvor sympotmer på hjertesvikt kan oppstå.
Trykkforandringer i atriet under hjertesyklusen
I figuren som betegner hjertesyklusen over vil du kunne se tre små endringer i kurven som betegner atrietrykk. a-bølgen forårsakes av atriekontraksjon, og trykket i det venstre atrie øker da normalt sett med 7-8 mm/Hg. c-bølgen oppstår når ventriklene begynner å kontrahere, som kan forårsaker en viss tilbakestrømning av blod til atriene men også at hjerteklaffene presses tilbake mot atriet på grunnet økt trykk i ventrikkelen. v-bølgen oppstår ved slutten på av den ventrikulære kontraksjonen, og kommer av den trege flyten av blod inn i atriene fra venene mens hjerteklaffene er lukket. Så, når den ventrikulære kontraksjonen er over, åpnes hjerteklaffene som fører til at atrielt blod strømmer ned i ventriklene, og v-bølgen opphører.
Ventriklene
Fylling av ventriklene under diastolen
Under ventrikulær systole samles blod i høyre og venstre atrium på grunn av lukkede hjerteklaffer, som hindrer at blodet strømmer ned i ventriklene når hjertet trekker seg sammen. På den måten, når den ventrikulære systolen opphører og disse slapper av, vil hjerteklaffene åpnes som følge av økt trykk atriene, og blodet vil på ny strømme inn i ventriklene. Dette kan du lese av figuren over der kurven som betegner ventrikkelvolum øker. Den ventrikulære diastolen kan deles opp i 3 faser hvor fase 1) er når ventriklene fylles hurtig ved at blodet strømmer rett igjennom atriene og inn i ventriklene, 2) betegner en liten mengde blod som tømmes i atriene fra venene under diastolen og som fortesetter rett til ventriklene, og 3) når atriene kontraherer og skviser den siste dråpen med blod fra ned i ventriklene.
Tømming av ventriklene under systolen
- Isometrisk fase
- Umiddelbart etter at ventriklene er begynt å kontrahere øker ventrikkeltrykket drastisk, som igjen fører til at hjerteklaffen som skiller atriet og ventrikkelen lukkes. Deretter passerer .02-.03 sekunder som ventrikkelen bruker til å bygge opp nok til trykk til å åpne aorta- (til hovedpulsåren) og pulmonalklaffen (til lungene) som man finner i henholdsvis venstre og høyre hjertehalvdel. Disse .02-.03 sekundene kalles den isometriske/isovolumiske perioden, ettersom ventrikkelen kontraherer men til tross for dette pumpes blodet likevel ikke ut av hjertet.
- Ejeksjonsfasen
- Når trykket i venstre ventrikkel øker til over 80 mm/Hg åpnes aorta- og pulmonalklaffen. Denne ejeksjonsfasen er også tredelt, og 70% av blodet strømmer ut gjennom ventriklene under den første tredjedelen, mens de resterende 30% pumpes ut til slutt. Første tredjedel kalles derfor den hurtige ejeksjonsfasen, mens de to siste tredjedelene kalles den trege ejeksjonsfasen.
- Isometrisk avslapningsfase
- Avslutningsvis i systolen opphører ventrikulær kontraksjon som gjør at det intraventrikulære trykket reduseres. Det økte trykket i de nå oppblåste, store arteriene som nettopp er fyllt med blod fører til at disse presser blodet tilbake mot ventriklene, som i tur fører til aortaklaffen og pulomnalklaffen lukkes. De neste .03-.06 sekundene slapper ventriklene av, og denne periodene kalles isometrisk avslapningsfase, og i denne fasen synker trykket i ventriklene raskt til sine diastoliske nivå. Deretter åpnes hjerteklaffene og et nytt hjerteslag tar form.
Endediastolisk volum, endesystolisk volum og slagvolum
Under diastole, økes normalt det ventrikulære volumet med rundt 110-120 ml. Dette volumet kalles endediastolisk volum (EDV). Deretter, når ventriklene tømmes under systolen, synker volumet med rundt 70 ml, og dette kalles slagvolum (SV). Det som er igjen i hver ventrikkel, ca 40-50 ml, kalles endesystolisk volum (ESV). Ved kraftig kontraksjon kan ESV reduseres til så lite som 10-20 ml. På den andre siden, når store mengder blod strømmer inn i ventriklene under diastolen kan EDV bli så stort som 150-180 ml i et friskt hjerte. Ved å øke EDV og samtidig redusere ESV kan SV mer enn fordobles som for eksempel under fysisk aktivitet.
Hjerteklaffene
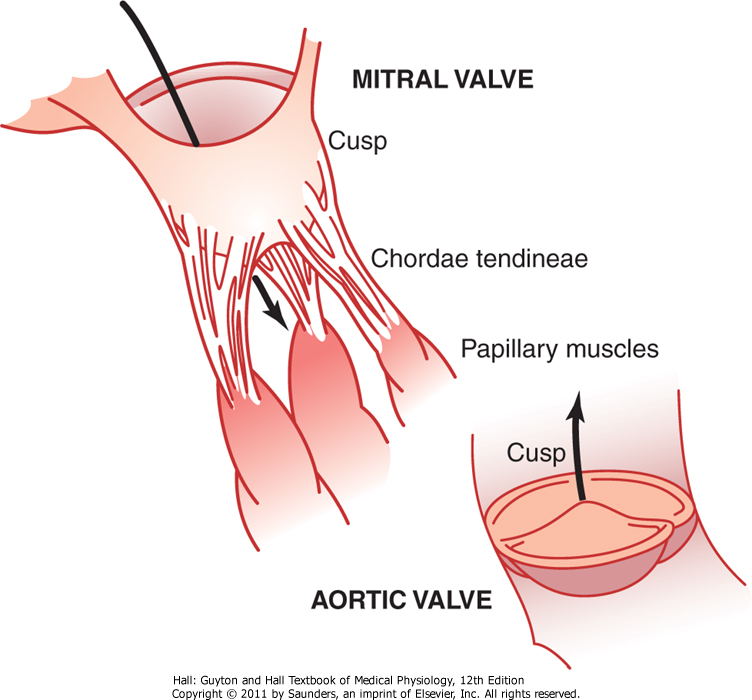
De atrioventrikulære klaffene
De atrioventrikulære klaffene (AV-klaffene) har jeg til nå i teksten kun betegnet som “hjerteklaffene”, og er klaffene som skiller atriene og ventriklene. Disse kalles også for trikuspidalklaffen (høyre hjertehalvdel) og mitralklaffen (venstre hjertehalvdel). AV-klaffene forhindrer tilbakestrømning av blod fra ventriklene til atriene under systolen. De lukkes når en tilbakegående trykkgradient presser blodet bakover og åpnes når det motsatte skjer.
Papillærmusklene
Papillærmusklene er muskler som er festet til AV-klaffene, og kontraherer samtidig med ventriklene. Til tross for at man skulle tro at disse musklene hjalp til med lukkingen av AV-klaffene, er det ikke det som skjer. Når ventriklene kontraherer, buler AV-klaffene inn mot sine respektive atrier, og det er papillærmusklene som holder disse igjen slik at de ikke buler for mye inn og begynner å lekke, som kan skje dersom dersom det skulle oppstå rifter eller skader i denne muskulaturen.
De semilunære klaffene
De semilunære klaffene omfatter aorta- og pulmonalklaffen. Disse halvmåneformede (derav navnet) klaffene skiller høyre og venstre ventrikkel fra henholdsvis pulmonalarterien og aorta, og forhindrer tilbakestrømning av blod til ventriklene under diastolen og ved systolens slutt når trykket i disse arteriene er høyt.
Blodtrykket
Når venstre ventrikkel kontraherer øker trykket inntil aortaklaffen åpnes. Når aortaklaffen åpnes avtar trykkøkningen, ettersom blodet pumpes ut av hjertet, inn i aorta, og ut i perifere vev. Når blodet kommer inn i de perifere arteriene strekkes åreveggene og trykket øker til rundt 120 mm/Hg (systolisk trykk). Når systolen avtar og aortaklaffen lukkes, opprettholdes arterietrykket grunnet elastisiteten i åreveggene. Dersom du tar en titt i figuren av hjertesyklusen vil du se dette beskrevet i kurven som betegner aortatrykket. Du vil også kunne se en kortvarig stigning i trykket som oppstår som følge av en liten men plutselig tilbakestrømning av blod rett før klaffen lukkes. Denne kortvarige trykkøkningen opphører like plutselig som den oppstår når aortaklaffen er lukket.
Etter at aortaklaffen er lukket synker trykket sakte men sikkert gjennom diastolen. Dette skjer fordi blodet som er kommet ut i de store arteriene flyter kontinuerlig gjennom de perifere årene og deretter tilbake til venene. Før ventrikkelen kontraherer igjen synker aortatrykket vanligvis til rundt 80 mm/Hg (diastolisk trykk), som er rundt 2/3-deler av trykket under systolen (120 mm/Hg). Det er dette legen måler når du sjekker blodtrykket, og tallene som er nevnt tilsvarer et normalt blodtrykk på 120/80.
Denne artikkelen er basert på kapittelet om hjertet i Guyton & Halls Textbook on Medical Physiology.